ძალიან მნიშვნელოვანია იმის გაგება, თუ როგორ ხდება ღვიძლში ქოლესტერინის სინთეზი. თუ ამ საკითხს დეტალურად შეისწავლით, მაშინვე გაირკვევა, რა კავშირი აქვს ღვიძლს ამ ორგანულ ნაერთთან. მაგრამ ჯერ უნდა გახსოვდეთ, რომ ნივთიერებას ასევე აქვს სახელი, რომელსაც ასევე ხშირად იყენებენ, კერძოდ, ქოლესტერინს.
როგორც ზემოთ უკვე აღვნიშნეთ, ეს ნივთიერება არის ორგანული ნაერთი და გვხვდება ყველა ცოცხალ ორგანიზმში. ის ლიპიდების განუყოფელი ნაწილია.
ყველაზე მაღალი კონცენტრაცია აღინიშნება ცხოველური წარმოშობის პროდუქტებში. მაგრამ მცენარეულ პროდუქტებში ამ ნაერთის მხოლოდ მცირე ნაწილია.
ასევე მნიშვნელოვანია აღინიშნოს ის ფაქტი, რომ საკვებთან ერთად ქოლესტერინის მთლიანი რაოდენობით მხოლოდ 20 პროცენტი შემოდის, დანარჩენი 80 პროცენტი ორგანიზმს დამოუკიდებლად აწარმოებს. სხვათა შორის, მთლიანი სინთეზირებული ნივთიერებისგან მხოლოდ 50% წარმოიქმნება ღვიძლში. ეს ხდება უჯრედულ დონეზე, დანარჩენი 30% წარმოებულია ნაწლავებში და კანში.
ადამიანის სხეული შეიცავს ამ კომპონენტის რამდენიმე ტიპს. ამავე დროს, უნდა აღინიშნოს, რომ ეს არის ჰემატოპოეზის სისტემა, რომელიც გაჯერებულია ამ ნივთიერებით. სისხლში ქოლესტერინი წარმოადგენს ცილის კომპლექსური ნაერთების ნაწილს, ასეთ კომპლექსებს უწოდებენ ლიპოპროტეინებს.
კომპლექსები შეიძლება იყოს ორი ტიპის:
- HDL - აქვთ ძალიან მაღალი სიმკვრივე, მათ უწოდებენ კარგს;
- LDL - აქვთ დაბალი სიმკვრივე, ამ ნივთიერებებს უწოდებენ ცუდი.
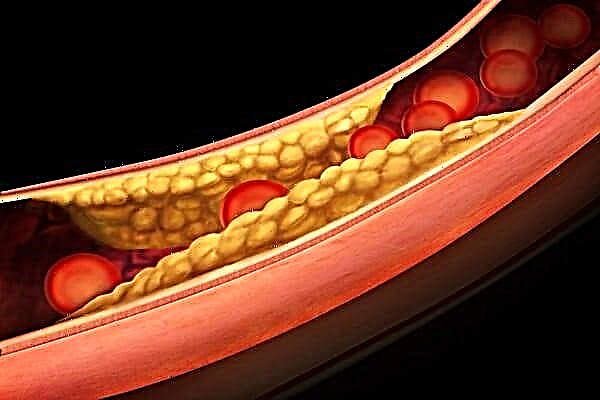
ეს არის მეორე ტიპი, რომელიც საფრთხეს უქმნის ადამიანისთვის. მას შემდეგ, რაც ისინი დაირბიან, რაც ნივთიერების კრისტალებისგან შედგება, ისინი იწყებენ დაგროვებას სისხლის მიმოქცევის სისტემის სისხლძარღვების კედლებზე დაფების სახით, რომლებიც პასუხისმგებელნი არიან სისხლის გადატანას. შედეგად, ეს პროცესი ხდება ისეთი პათოლოგიების ორგანიზმში განვითარების მიზეზი, როგორიცაა ათეროსკლეროზი.
ათეროსკლეროზის პროგრესირება იწვევს მრავალი სერიოზული დაავადების განვითარებას.
კავშირის ძირითადი მახასიათებლები
როგორც ზემოთ აღინიშნა, ეს ნივთიერება შეიძლება სასარგებლო იყოს ადამიანისთვის, რა თქმა უნდა, მხოლოდ იმ შემთხვევაში, თუ HDL- ზე ვსაუბრობთ.
ამის საფუძველზე ცხადი ხდება, რომ შეცდომაა იმის მტკიცება, რომ ქოლესტერინი აბსოლუტურად საზიანოა ადამიანისთვის.
ქოლესტერინი არის ბიოლოგიურად აქტიური კომპონენტი:
- მონაწილეობს სქესის ჰორმონების სინთეზში;
- უზრუნველყოფს ტვინში სეროტონინის რეცეპტორების ნორმალურ ფუნქციონირებას;
- არის ნაღვლის მთავარი კომპონენტი, ასევე D ვიტამინი, რომელიც პასუხისმგებელია ცხიმების შეწოვაში;
- თავისუფალი რადიკალების გავლენის ქვეშ ხელს უშლის უჯრედშორისი სტრუქტურების განადგურების პროცესს.
მაგრამ დადებით თვისებებსთან ერთად, ნივთიერებას შეუძლია ზიანი მიაყენოს ადამიანის ჯანმრთელობას. მაგალითად, LDL შეიძლება გამოიწვიოს სერიოზული დაავადებების განვითარება, პირველ რიგში, ათეროსკლეროზის განვითარებაში.
ღვიძლში, ბიოკომპონენტი სინთეზირდება HMG რედუტაზის გავლენის ქვეშ. ეს არის მთავარი ფერმენტი, რომელიც მონაწილეობს ბიოსინთეზში. სინთეზის დათრგუნვა ხდება უარყოფითი გამოხმაურების გავლენის ქვეშ.
ნივთიერებათა ღვიძლში ნივთიერების სინთეზის პროცესს აქვს ინვერსიული კავშირი ნაერთის იმ დოზასთან, რომელიც ადამიანის სხეულში შემოდის საკვებით.
კიდევ უფრო მარტივი, ეს პროცესი ამ გზით არის აღწერილი. ღვიძლი დამოუკიდებლად არეგულირებს ქოლესტერინის დონეს. რაც უფრო მეტ ადამიანი მოიხმარს ამ კომპონენტის შემცველ საკვებს, მით ნაკლები ნივთიერება იწარმოება ორგანოს უჯრედებში და თუ გავითვალისწინებთ, რომ ცხიმები მოიხმარენ მის შემცველ პროდუქტებთან ერთად, ეს მარეგულირებელი პროცესი ძალიან მნიშვნელოვანია.
მატერიის სინთეზის თვისებები
ნორმალური ჯანმრთელი მოზრდილები სინთეზს უწევენ HDL– ს დაახლოებით 1 გრ / დღეში და მოიხმარენ დაახლოებით 0.3 გ დღეში.
 სისხლში ქოლესტერინის შედარებით მუდმივ დონეს აქვს ასეთი მნიშვნელობა - 150-200 მგ / დლ. ინახება ძირითადად დენოვოს სინთეზის დონის კონტროლით.
სისხლში ქოლესტერინის შედარებით მუდმივ დონეს აქვს ასეთი მნიშვნელობა - 150-200 მგ / დლ. ინახება ძირითადად დენოვოს სინთეზის დონის კონტროლით.
მნიშვნელოვანია აღინიშნოს, რომ ენდოგენური წარმოშობის HDL და LDL სინთეზი ნაწილობრივ რეგულირდება დიეტებით.
ქოლესტერინი, როგორც საკვებისგან, ასევე ღვიძლში სინთეზირებული, გამოიყენება მემბრანების ფორმირებაში, სტეროიდული ჰორმონების და ნაღვლის მჟავების სინთეზში. ნივთიერების უდიდესი წილი გამოიყენება ნაღვლის მჟავების სინთეზში.
უჯრედების მიერ HDL და LDL- ის მიღება სტაბილურ დონეზე შენარჩუნებულია სამი სხვადასხვა მექანიზმით:
- HMGR– ის საქმიანობის რეგულირება
- უჯრედშიდა თავისუფალი ქოლესტერინის ჭარბი რეგულირება O-acyltransferase სტეროლის, SOAT1 და SOAT2– ის მოქმედებით SOAT2– ით, რაც ღვიძლში ძირითადად აქტიური კომპონენტია. ამ ფერმენტებისთვის საწყისი დანიშნულება იყო ACAT აცილ-CoA- სთვის: აცილტრანსფერაზას ქოლესტერინი. ფერმენტები ACAT, ACAT1 და ACAT2 არის აცეტილ CoA აცეტილტრანსფერაზები 1 და 2.
- პლაზმაში ქოლესტერინის დონის კონტროლის გზით LDL შუამავლობით რეცეპტორების შეყვანა და HDL შუამავლობით საპირისპირო ტრანსპორტი.
HMGR საქმიანობის რეგულირება არის LDL და HDL ბიოსინთეზის დონის კონტროლის ძირითადი საშუალება.
ფერმენტი კონტროლდება ოთხი განსხვავებული მექანიზმით:
- უკუკავშირის ინჰიბიცია;
- გენის გამოხატვის კონტროლი;
- ფერმენტების დეგრადაციის მაჩვენებელი;
- ფოსფორილირება-დეფოსფორილირება.
პირველი სამი მაკონტროლებელი მექანიზმი მოქმედებს უშუალოდ თავად ნივთიერებაზე. ქოლესტერინი მოქმედებს, როგორც უკუკავშირი ინჰიბიტორი ადრეული HMGR და ასევე იწვევს ფერმენტის სწრაფ დეგრადაციას. ეს უკანასკნელი HMGR– ის პოლიუბიკტილაციის შედეგია და მისი დეგრადაცია პროტეინოზულში. ეს უნარი HMGR SSD სტეროლის მგრძნობიარე დომენის შედეგია.
გარდა ამისა, როდესაც ქოლესტერინი ჭარბი რაოდენობითაა, HMGR- სთვის mRNA- ს რაოდენობა მცირდება გენის გამოხატვის შედეგად.
სინთეზში მონაწილე ფერმენტები
 თუ ეგზოგენური კომპონენტი რეგულირდება კოვალენტური მოდიფიკაციის გზით, ეს პროცესი განხორციელდება ფოსფორილირებისა და დეფოსფორილირების შედეგად.
თუ ეგზოგენური კომპონენტი რეგულირდება კოვალენტური მოდიფიკაციის გზით, ეს პროცესი განხორციელდება ფოსფორილირებისა და დეფოსფორილირების შედეგად.
ფერმენტი ყველაზე აქტიურია არამოდიფიცირებული ფორმით. ფერმენტის ფოსფორილირება ამცირებს მის აქტივობას.
HMGR ფოსფორილირდება AMP- ით გააქტიურებული ცილა კინაზით, AMPK. თავად AMPK გააქტიურებულია ფოსფორილირებით.
AMPK ფოსფორილირება ხდება მინიმუმ ორი ფერმენტის კატალიზებით, კერძოდ:
- პირველადი კინაზა, რომელიც პასუხისმგებელია AMPK გააქტიურებაზე, არის LKB1 (ღვიძლის კინაზა B1). LKB1 პირველად იქნა გამოვლენილი, როგორც გენი ადამიანებში, რომლებსაც აქვთ აუტოსომური დომინანტური მუტაცია პუც-ჯეგერების სინდრომში, PJS. LKB1 ასევე გვხვდება, რომ მუტანტია ფილტვის ადენოკარცინომაში.
- მეორე ფოსფორილირებადი ფერმენტი AMPK არის კალმოდულინზე დამოკიდებული ცილა კინაზა კინაზა ბეტა (CaMKKβ). CaMKKβ იწვევს AMPK ფოსფორილირებას კუნთების შეკუმშვის შედეგად უჯრედშიდა Ca2 + - ით გაზრდის საპასუხოდ.
HMGR– ის რეგულირება კოვალენტური მოდიფიკაციით საშუალებას იძლევა HDL წარმოება. HMGR ყველაზე აქტიურია დეფოსფორილირებულ მდგომარეობაში. ფოსფორილირება (Ser872) კატალიზირდება AMP– აქტიური ცილის კინაზის (AMPK) ფერმენტის საშუალებით, რომლის მოქმედება ასევე რეგულირდება ფოსფორილირებით.
AMPK ფოსფორილირება შეიძლება მოხდეს მინიმუმ ორი ფერმენტის გამო:
- LKB1;
- CaMKKβ.
HMGR– ის დეფოსფორილირება, მისი უფრო აქტიურ მდგომარეობაში დაბრუნება, ხორციელდება 2A ოჯახის ცილოვანი ფოსფატაზების მოქმედებით. ეს თანმიმდევრობა საშუალებას გაძლევთ გააკონტროლოთ HDL- ის წარმოება.
რა გავლენას ახდენს ქოლესტერინის ტიპზე?
 ფუნქციური PP2A არსებობს ორი განსხვავებული კატალიზური იზოფორმის მიერ, რომლებიც კოდირებულია ორი გენის მიერ, რომლებიც იდენტიფიცირებულია PPP2CA და PPP2CB. PP2A- ს ორი ძირითადი იზოფორმია ჰეტეროდიმერული ბირთვიანი ფერმენტი და ჰეტეროტრიმერული ჰოლოენზიმი.
ფუნქციური PP2A არსებობს ორი განსხვავებული კატალიზური იზოფორმის მიერ, რომლებიც კოდირებულია ორი გენის მიერ, რომლებიც იდენტიფიცირებულია PPP2CA და PPP2CB. PP2A- ს ორი ძირითადი იზოფორმია ჰეტეროდიმერული ბირთვიანი ფერმენტი და ჰეტეროტრიმერული ჰოლოენზიმი.
მთავარი PP2A ფერმენტი შედგება სკაფონის ქვესტატისგან (თავდაპირველად უწოდებენ A ქვენატს) და კატალიტიკური ქვედანაყოფი (C ქვემყოფი). კატალიტიკური α ქვესისტემა კოდირდება PPP2CA გენის მიერ, ხოლო კატალიზური β ქვესისტემა კოდირდება PPP2CB გენის მიერ.
Α სკაფონის ქვესტრუქტურა კოდირდება PPP2R1A გენის და β ქვესიტენის მიერ PPP2R1B გენის მიერ. მთავარი ფერმენტი, PP2A, ურთიერთქმედებს ცვლადი მარეგულირებელი ქვედანაყოფით, რომ შეიკრიბოს ჰოლოენზიმი.
PP2A საკონტროლო ქვედანაყოფი მოიცავს ოთხ ოჯახს (თავდაპირველად უწოდებენ B- ქვეგანყოფილებებს), რომელთაგან თითოეული შედგება სხვადასხვა გენების მიერ დაშიფრული რამდენიმე იზოფორმისგან.
ამჟამად, PP2A B. მარეგულირებელი ქვედანაყოფისთვის არსებობს 15 სხვადასხვა გენი. PP2A მარეგულირებელი ქვედანაყოფის ძირითადი ფუნქციაა PP2A- ს კატალიზური ქვედანაყოფების ფოსფატაზური აქტივობის ფოსფორილირებული სუბსტრატის ციტირება.
PPP2R არის PP2A 15 სხვადასხვა მარეგულირებელი ქვედანაყოფიდან ერთ-ერთი. ისეთი ჰორმონები, როგორიცაა გლუკაგონი და ადრენალინი, უარყოფითად მოქმედებს ქოლესტერინის ბიოსინთეზზე, ფერმენტების PP2A ოჯახის სპეციფიური მარეგულირებელი ქვედანაყოფების აქტივობის გაზრდით.
PP2A (PPP2R) მარეგულირებელი ქვედანაყოფის PKA- შუამავლობით ფოსფორილირება იწვევს PP2A– ს განთავისუფლებას HMGR– დან, მისი დეფოსფორილირების თავიდან ასაცილებლად. გლუკაგონისა და ადრენალინის ეფექტების საწინააღმდეგოდ, ინსულინი ასტიმულირებს ფოსფატების მოცილებას და ამით ზრდის HMGR– ის მოქმედებას.
HMGR– ის დამატებითი რეგულირება ხდება ქოლესტერინით უკუკავშირის ინჰიბიციის გზით, აგრეთვე მისი სინთეზის რეგულირება უჯრედშორისი ქოლესტერინის და სტეროლის დონის ამაღლებით.
ეს უკანასკნელი ფენომენი ასოცირდება ტრანსკრიფციის ფაქტორთან SREBP.
როგორ ხდება პროცესი ადამიანის სხეულში?
 HMGR საქმიანობას დამატებით აკონტროლებს AMP– ით სიგნალიზაცია. CAMP– ის ზრდა იწვევს cAMP– ზე დამოკიდებული ცილოვანი კინაზის, PKA– ს გააქტიურებას. HMGR რეგულირების კონტექსტში, PKA ფოსფორილირებს მარეგულირებელ ქვედანაყოფს, რაც განაპირობებს PP2A- სგან განთავისუფლებას HMGR– დან. ეს ხელს უშლის PP2A ფოსფატებს HMGR– ის ამოღებას, ხელს უშლის მის რეაქტირებას.
HMGR საქმიანობას დამატებით აკონტროლებს AMP– ით სიგნალიზაცია. CAMP– ის ზრდა იწვევს cAMP– ზე დამოკიდებული ცილოვანი კინაზის, PKA– ს გააქტიურებას. HMGR რეგულირების კონტექსტში, PKA ფოსფორილირებს მარეგულირებელ ქვედანაყოფს, რაც განაპირობებს PP2A- სგან განთავისუფლებას HMGR– დან. ეს ხელს უშლის PP2A ფოსფატებს HMGR– ის ამოღებას, ხელს უშლის მის რეაქტირებას.
მარეგულირებელი ცილოვანი ფოსფატაზის ქვედანაყოფების დიდი ოჯახი არეგულირებს ან / და აფერხებს მრავალ ფოსფატაზების მოქმედებას, მათ შორისაა PP1, PP2A და PP2C ოჯახების წევრები. PP2A ფოსფატაზების გარდა, რომლებიც ფოსფატებს აცილებენ AMPK და HMGR, ცილის ფოსფატაზას 2C ოჯახის ფოსფატაზები (PP2C) ასევე ამოიღებენ ფოსფატებს AMPK- დან.
როდესაც ეს მარეგულირებელი ქვედანაყოფი ფოსფორილატს PKA იწვევს, შეკრული ფოსფატაზების მოქმედება კლებულობს, რის შედეგადაც AMPK დარჩება ფოსფორილირებულ და აქტიურ მდგომარეობაში, ხოლო HMGR ფოსფორილირებულ და არააქტიურ მდგომარეობაში. სტიმულის მოცილების გამო, რასაც იწვევს cAMP წარმოების ზრდა, მცირდება ფოსფორილირების დონე და იზრდება დეფოსფორილირების დონე. საბოლოო შედეგი არის HMGR საქმიანობის უფრო მაღალ დონეზე დაბრუნება. თავის მხრივ, ინსულინი იწვევს cAMP– ის დაქვეითებას, რაც, თავის მხრივ, ააქტიურებს სინთეზს. საბოლოო შედეგი არის HMGR საქმიანობის უფრო მაღალ დონეზე დაბრუნება.
თავის მხრივ, ინსულინი იწვევს კამპუსის შემცირებას, რაც, თავის მხრივ, ააქტიურებს ქოლესტერინის სინთეზს. საბოლოო შედეგი არის HMGR საქმიანობის უფრო მაღალ დონეზე დაბრუნება. ინსულინი იწვევს კამპუსის შემცირებას, რაც, თავის მხრივ, შეიძლება გამოყენებულ იქნას სინთეზის პროცესის გასაძლიერებლად.
ინსულინის სტიმულირებისა და გლუკაგონის ინჰიბირების უნარი, HMGR– ის მოქმედება შეესაბამება ამ ჰორმონების გავლენას სხვა მეტაბოლურ მეტაბოლურ პროცესებზე. ამ ორი ჰორმონის მთავარი ფუნქციაა უჯრედებზე წვდომის კონტროლი და ენერგიის ტრანსპორტირება.
HMGR– ის მოქმედების გრძელვადიანი კონტროლი ხორციელდება ძირითადად ფერმენტის სინთეზის და დეგრადაციის კონტროლით. როდესაც ქოლესტერინის დონე მაღალია, HMGR გენის გამოხატვის დონე მცირდება, და პირიქით, უფრო დაბალი დონე ააქტიურებს გენის გამოხატვას.
ქოლესტერინის შესახებ ინფორმაცია მოცემულია ამ სტატიაში ვიდეოში.